Endocytic trafficking
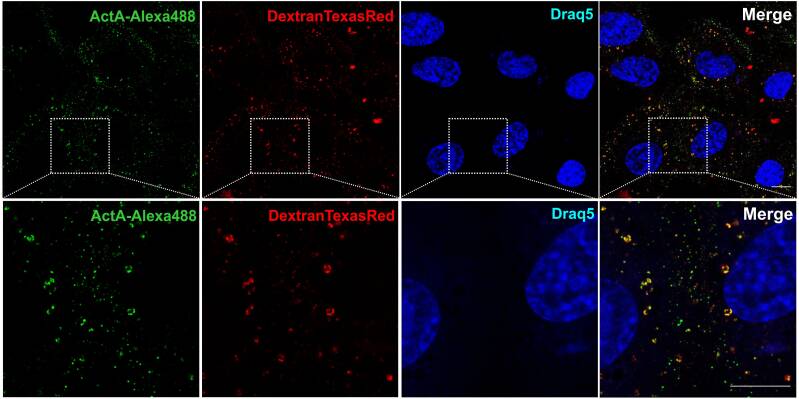
Trafficking Pathway for Activin A in human embryonic stem and differentiated Cells
Ligand/receptor complexes formed at the plasma membrane are internalised via various endocytic pathways that influence the ultimate signalling output by regulating the complex’s selection of interaction partners along the trafficking route. We have shown that in differentiated cells Activin A/receptor complexes are internalised via Clathrin-mediated endocytosis (CME) and macropinocytosis (MP), whereas in human embryonic stem cells (hESCs) internalisation occurred via CME. We further show that hESCs are devoid of MP, which becomes functional upon differentiation towards endothelial cells through mesoderm mediators. Our results reveal, for the first time, that MP is an internalisation route of Activin A in differentiated cells, which is not active in hESCs and is induced as cells differentiate.
Kostopoulou N., Bellou S., Bagli E., Markou M., Kostaras E., Hyvönen M., Kalaidzidis Y., Papadopoulos A., Chalmantzi V., Kyrkou A., Panopoulou E., Fotsis T., Murphy C.; Embryonic stem cells are devoid of macropinocytosis, a trafficking pathway for activin A in differentiated cells. J Cell Sci 1 July 2021; 134 (13): jcs246892. doi: https://doi.org/10.1242/jcs.246892

ERBIN is a new SARA-interacting protein: competition between SARA and SMAD2 and SMAD3 for binding to ERBIN
SARA, an early endosomal protein, plays a key role in TGFβ signalling, as it presents SMAD2 and SMAD3 for phosphorylation by the activated TGFβ receptors. We have shown that ERBIN is a new SARA-interacting protein that can be recruited by SARA to early endosomes. ERBIN was recently shown to bind and segregate phosphorylated SMAD2 and SMAD3 (SMAD2/3) in the cytoplasm, thereby inhibiting SMAD2/3-dependent transcription. SARA binds to ERBIN using a new domain, which we have called the ERBID (ERBIN-binding domain), whereas ERBIN binds to SARA using a domain (amino acids 1208-1265) that also interacts with SMAD2 and SMAD3, which we have called the SSID (SARA- and SMAD-interacting domain). We have additionally shown that SARA competes with SMAD2/3 for binding to ERBIN. In agreement, overexpression of SARA or the ERBID peptide reverses the inhibitory effect of ERBIN on SMAD2/3-dependent transcription. Taken together, these data suggest that the response of cells to TGFβ and activin A can be influenced by the relative concentrations of SARA, ERBIN and SMAD2/3.
Sflomos G, Kostaras E, Panopoulou E, Pappas N, Kyrkou A, Politou AS, Fotsis T, Murphy C. ERBIN is a new SARA-interacting protein: competition between SARA and SMAD2 and SMAD3 for binding to ERBIN. J Cell Sci. 2011 Oct 1;124(Pt 19):3209-22. doi: 10.1242/jcs.062307. Epub 2011 Aug 30. PMID: 21878490.
SARA and RNF11 at the crossroads of EGFR signaling and trafficking
The classical view that endocytosis serves only for growth factor receptor degradation and signaling termination has been challenged by an increasing number of reports showing that various growth factor receptors, such as epidermal growth factor receptor (EGFR), continue to activate downstream signaling molecules en route to lysosomes prior to their degradation. Moreover, the trafficking route that the ligand-receptor complexes follow to enter the cell is mutually interconnected with the final signaling output. Endosomal resident effector proteins are compartmentalized and regulate the signaling and trafficking of the ligand-bound receptor complexes. Smad anchor for receptor activation (SARA) is an early endosomal protein facilitating TGF-β signaling cascade. Even though SARA was identified as an adaptor protein that regulates SMAD2 activation and TGF-β signal propagation, an increasing number of reports in various systems describe SARA as a trafficking regulator. SARA has been shown to interact with the E3 ubiquitin ligase RNF11 (RING finger protein 11) and members of the ESCRT-0 (endosomal sorting complex required for transport) complex functionally participating in the degradation of EGFR.
Kostaras E, Pedersen NM, Stenmark H, Fotsis T, Murphy C. SARA and RNF11 at the crossroads of EGFR signaling and trafficking. Methods Enzymol. 2014;535:225-47. doi: 10.1016/B978-0-12-397925-4.00014-6. PMID: 24377927.


Early endosomal regulation of Smad-dependent signaling in endothelial cells
Transforming growth factor beta (TGFbeta) receptors require SARA for phosphorylation of the downstream transducing Smad proteins. SARA, a FYVE finger protein, binds to membrane lipids suggesting that activated receptors may interact with downstream signaling molecules at discrete endocytic locations. We revealed a critical role for the early endocytic compartment in regulating Smad-dependent signaling. Not only is SARA localized on early endosomes, but also its minimal FYVE finger sequence is sufficient for early endosomal targeting. Expression of a SARA mutant protein lacking the FYVE finger inhibits downstream activin A signaling in endothelial cells. Moreover, a dominant-negative mutant of Rab5, a crucial protein for early endosome dynamics, causes phosphorylation and nuclear translocation of Smads leading to constitutive (i.e. ligand independent) transcriptional activation of a Smad-dependent promoter in endothelial cells. As inhibition of endocytosis using the K44A negative mutant of dynamin and RN-tre did not lead to activation of Smad-dependent transcription, the effects of the dominant-negative Rab5 are likely to be a consequence of altered membrane trafficking of constitutively formed TGFbeta/activin type I/II receptor complexes at the level of early endosomes. The results suggest an important interconnection between early endosomal dynamics and TGFbeta/activin signal transduction pathways.
Panopoulou E, Gillooly DJ, Wrana JL, Zerial M, Stenmark H, Murphy C, Fotsis T. Early endosomal regulation of Smad-dependent signaling in endothelial cells. J Biol Chem. 2002 May 17;277(20):18046-52. doi: 10.1074/jbc.M107983200. Epub 2002 Mar 4. PMID: 11877415.
Figure legend: Intracellular localization of SARA in endothelial cells. BBCE cells transfected with FLAG-tagged SARA were processed for immunofluorescence using anti-FLAG antibody (a, e, g, and j). EEA1 was stained by a specific antibody/fluorescein isothiocyanate-labeled secondary antibody complex (b), GFP-Rab5Q79L was detected by visualizing GFP directly (d), human transferrin receptors were labeled by fluorescent Alexa-transferrin uptake (h), and late endosomes/lysosomes were localized by Lysotracker (k). Overlays are shown in c, f,i, and l. Size bars are 10 μm.